
Contribution of satellite cells to IGF-I induced hypertrophy of skeletal muscle
O aumento do tamanho do músculo esquelético – hipertrofia muscular – ocorre em resposta a demandas funcionais. Alguns modelos animais foram criados, como hipertrofia induzida por estiramento, ablação de musculatura sinergista e infusão ou super-expressão de IGF-1. Como o músculo esquelético é um tecido pós-mitótico, mudanças no volume do mesmo dependem de uma fonte externa de células mitoticamente ativas. As células que desempenham este papel são as células-satélites, situadas entre o sarcolema e a lâmina basal da fibra muscular. Elas podem se diferenciar em mionúcleos, mantendo a relação núcleos/sarcoplasma (dominímio mionuclear)constante mesmo durante o processo de hipertrofia. Além disso, quando o músculo esquelético aumenta em tamanho, é predominantemente devido à ativação, proliferação e fusão das células satélites às fibras pré-existentes, como acontece após lesão por aumento de tensão (p. ex. treinamento de força).
Os autores do presente trabalho observaram anteriormente que a expressão aumentada de IGF-1 no músculo promoveu hipertrofia significativa e aumento concomitante de força. Observaram também um aumento de fibras com núcleos centralizados, através de análise histológica. Isto é indicativo de fusão recente de mioblastos, que ocorre após a proliferação das células satélites. Desta forma, o objetivo do trabalho é determinar a contribuição das células satélites na hipertrofia induzida pelo IGF-1.
Para tanto, os autores utilizaram camundongos C57 divididos em três grupos: a)submetidos à radiação gama (X-ray : o que inibe a proliferação das células satélites); b) que super-expressam IGF-1 muscular (+IGF-1); c) com super-expressão do IGF-1 e submetidos à radiação gama.
Os resultados mostraram que os músculos +IGF-1 demonstraram maior massa muscular, quando comparados à para controle não tratada. Além disso, os músculos +X-ray tiveram uma massa reduzida, demonstrando que as células satélites são necessárias para o crescimento normal do músculo. Por fim, o aumento da massa muscular observada nos animais +IGF-1 foi prevenida pela radiação gama (+X-ray/IGF-1), conforme mostra a figura abaixo.
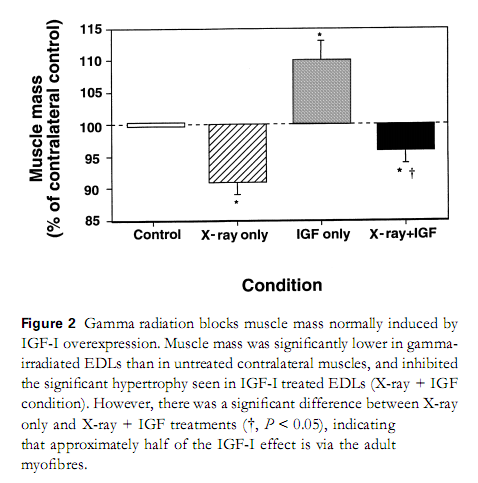
Em conclusão, o estudo de Barton-Davis et al (1999) nos mostra que as células satélites desempenham um papel crítico na hipertrofia induzida pela super-expressão do IGF-1. Entretanto, nos indica também que o IGF-1 exerce parte de seus efeitos hipertróficos via ação direta nas fibras musculares diferenciadas.
Referência: Barton-Davis et al. Contribution of satellite cells to IGF-I induced hypertrophy of skeletal muscle. Acta Physiol Scand 1999, 167, 301-305.