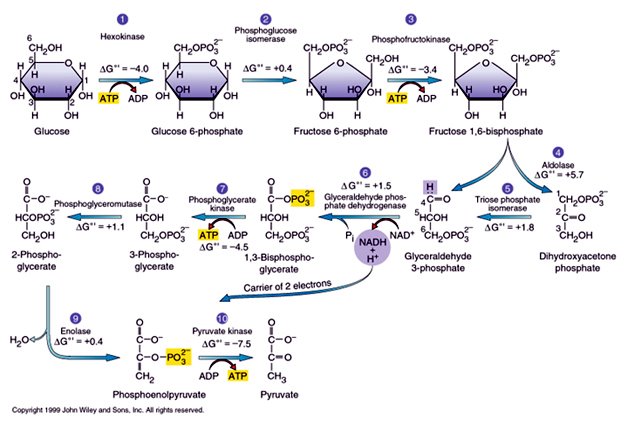
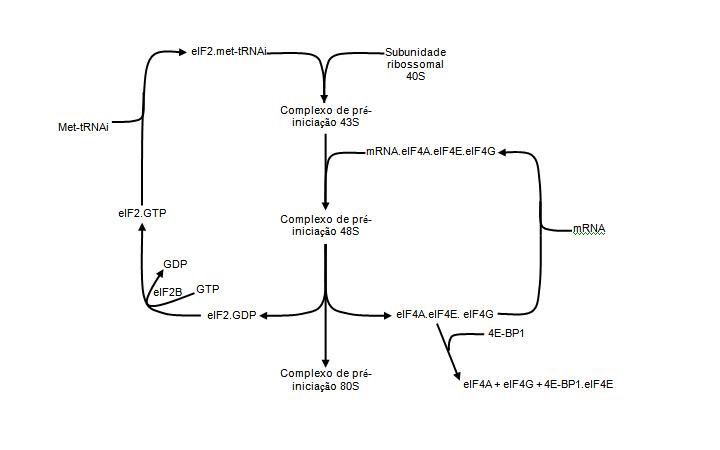
O CreaT é uma proteína integral de membrana, com 12 domínios trans-membrana e aproximadamente 70,5 kDa, apresentando cadeia de 635 aminoácidos (SNOW e MURPHY, 2001). Sua estrutura é muito similar às dos transportadores de dopamina, GABA, taurina e noradrenalina, todos dependentes de Na+/Cl-. Três sítios de glicosilação foram encontrados ao longo da cadeia do CreaT, sendo dois nas alças extracelulares entre os domínios transmembrana 3 e 4 e um entre o 11 e 12 (SALTARELLI et al., 1996; SORA et al., 1994). Além disso, sítios de fosforilação foram identificados na terminação amino e carboxílica, bem como em alças intracelulares, totalizando cinco locais de fosforilação (SALTARELLI et al., 1996; SORA et al., 1994). Os estados de glicosilação e fosforilação da proteína são possíveis moduladores da atividade do CreaT e, conseqüentemente, da taxa de captação de Cr pela célula (NASH et al., 1994; ZHAO et al., 2002).
O CreaT foi identificado em diversos tecidos, como o músculo esquelético, cérebro, miocárdio, rins, testículos, fígado, pulmões, enterócitos, retina e eritrócitos (GUIMBAL e KILIMANN, 1993; SNOW e MURPHY, 2001; SPEER, et al., 2004; TOSCO et al., 2004; NAKASHIMA et al., 2004). Quanto à localização celular do transportador, foi demonstrado, através de imagem por fluorescência e análise imunohistoquímica, que o CreaT está localizado juntamente com a isoforma α1 da Na+/K+ ATPase e a enzima citrato sintase, presentes, respectivamente, nas membranas plasmática e mitocondrial (SNOW e MURPHY, 2001). O transporte da Cr através deste transportador utiliza um sistema de co-transporte com Cl- e Na+, com estequeometria de 2 Na+ e 1 Cl- por molécula de Cr transportada (GARCIA-DELGADO et al., 2001 e PERAL et al., 2002), utilizando a energia do gradiente eletroquímico do Na+, gerado pela Na+/K+ ATPase (GUERRERO-ONTIVEROS e WALLIMANN, 1998).
Mais estudos sobre a regulação da expressão e atividade do CreaT possibilitará a melhor compreensão dos efeitos da suplementação de creatina e, especialmente, auxiliará do tratamento de algumas patologias que envolvem o sistema, como doenças neurodegenerativas, miopatias, além das moléstias que levam a intensa perda de massa e função musculares.
Por Lucas Guimarães Ferreira