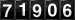Muito falamos em síntese de proteínas. Mas como este evento celular acontece?
Este vídeo explica de forma bem clara e didática.
[youtube http://www.youtube.com/watch?v=-ygpqVr7_xs&hl=en&fs=1&rel=0&color1=0x2b405b&color2=0x6b8ab6&border=1]
Muito falamos em síntese de proteínas. Mas como este evento celular acontece?
Este vídeo explica de forma bem clara e didática.
[youtube http://www.youtube.com/watch?v=-ygpqVr7_xs&hl=en&fs=1&rel=0&color1=0x2b405b&color2=0x6b8ab6&border=1]
Esmark e colaboradores realizaram um estudo no qual homens treinaram um protocolo de exercício de força por 12 semanas. Parte consumiu proteínas imediatamente após a sessão de treinamento e parte 2 horas após esta sessão. Após as 12 semanas, o grupo que fez a suplementação imediatamente após o exercício apresentou um maior ganho de massa muscular, com maior diâmetro das fibras musculares.
Levenhagen e colaboradores, por sua vez, verificaram a influência do tempo de suplementação, imediatamente ou 3 horas após exercício intenso em bicicleta, sobre a incorporação de aminoácidos no músculo. Mais uma vez, a suplementação imediatamente após o exercício mostrou-se mais eficaz.
Já Tipton e colaboradores demonstraram que o consumo de aminoácidos, imediatamente antes do exercício levou a uma maior síntese protéica muscular quando comparado ao consumo feito imediatamente após. Os autores atribuem este efeito à uma maior entrega de aminoácidos aos músculos, devido ao aumento do fluxo sanguíneo nos músculos que estão sendo exercitados.
Em contrapartida, um outro trabalho (Rasmussen et al., 2000) não encontrou diferença no balanço líquido de aminoácidos entre os que consumiram aminoácidos 1 ou 3 horas após o exercício.
Que lição podemos tirar destes trabalhos? A existência de um período ótimo para suplementação protéica é uma questão ainda especulativa, mas atrativa e provável. De forma prática, devemos realizar esta suplementação nos momentos que cercam a sessão de treinamento, ou seja, imediatamente antes, e/ou imediatamente após. Desta forma, promovemos um maior estímulo da síntese protéica muscular, possibilitando maiores ganhos de massa muscular.
Bons treinos!
Referências Bibliográficas
ESMARCK et al. Timing of postexercise protein intake is important for muscle hypertrophy with resistance training in elderly humans. J. Physiol. 535:301-11, 2001.
LEVENHAGEN et al. Postexercise nutrient intake timing in humans is critical to recovery of leg glucose protein homeostasis. Am. J. Physiol. Endocrinol. Metab. 280:E982-93, 2001.
TIPTON et al. Timing of aminoacid-carbohydrate ingestion alters anabolic response of muscle to resistance exercise. Am. J. Physiol. Endocrinol. Metab. 281: E197-206, 2001.
RASMUSSEN et al., An oral essential aminoacid-carbohydrate suplement enhances muscle protein anabolism after resistance exercise. J. Appl. Physiol. 88:386-92, 2000.
———————-
Lucas Guimarães
O sistema ubiquitina-proteassoma é o sistema proteolítico mais recentemente descoberto, sendo dependente de ATP. A vasta maioria das proteínas celulares é degradada através deste sistema (CAO et al., 2005). Este processo proteolítico envolve a participação de um complexo enzimático denominado proteassoma 26S, formado por 2 subunidades regulatórias (19S) e uma subunidade catalítica (20S). A subunidade 19S reconhece proteínas “marcadas” com uma cadeia de ubiquitina, um peptídeo de 76 aminoácidos (CIECHANOVER, 2005).
Três componentes enzimáticos são necessários para ligar a cadeia de ubiquitina nas proteínas destinadas à degradação: as enzimas E1 (ativadora de ubiquitina) e E2 (proteínas conjugadoras de Ub) preparam a Ub para conjugação, enquanto que as E3 (Ub-ligases), as enzimas-chave do processo, liga a Ub à proteína, que é então reconhecida pelo proteassoma 26S, um complexo proteolítico multi-unidades e multi-catalítico que degrada as proteínas ubiquitinadas em fragmentos menores (figura 3; CAO et al., 2005; CIECHANOVER, 2005).

Fig 3: sistema ubiquitina-proteassoma (adaptado de Jefferson et al.,2001).
Existe apenas uma proteína E1, algumas E2, e milhares de E3. Desta forma, esta última é o componente que confere especificidade ao sistema (CAO et al., 2005). Das diversas ubiquitina-ligases conhecidas, está bem estabelecido na literatura que atrogin-1 (MAFbx), MuRF-1 e E3α têm grande importância no processo de atrofia muscular. (LECKER et al., 1999a; 1999b; 2004; BODINE et al., 2001; GOMES et al., 2001, JONES et al., 2004). Verificou-se que E3α atua em conjunto com uma proteína conjugadora de ubiquitina, denominada E214k, que também tem sua expressão aumentada em situações de atrofia (WING & BANVILLE, 1994; LECKER et al., 1999a; 1999b).
A ubiquitinação das proteínas é um processo reversível. Enzimas desubiquitinadoras desempenham um importante papel na proteólise ubiquitina-dependente, catalisando a remoção da Ub das proteínas, livrando-as da degradação (KIM et al., 2003).
Foi demonstrado que o passo inicial para a proteólise via sistema ubiquitina-proteassoma no músculo esquelético parece ocorrer pela ação da caspase-3. Uma vez que o proteassoma não é capaz de clivar diretamente os complexos de actomiosina e as miofibrilas, verificou-se que esta caspase cliva estas estruturas em peptídeos menores, com cerca de 14 kDa, que são então ubiquitinados e, assim, degradados pelo proteassoma 26S (DU et al., 2004; 2005). Além disso, há evidências indicando que as calpaínas poderiam também participar da clivagem inicial do complexo de actomiosina (HUANG & FORSBERG, 1998; WILLIAMS et al., 1999).
Em condições que levam à atrofia muscular, desvernação, hipertireoidismo e sépse, a utilização de um inibidor de proteassoma (MG132) reduziu a proteólise muscular em 70%, 40-70% e 100%, respectivamente, demonstrando a importância do sistema ubiquitina-proteassoma em condições de intenso catabolismo protéico muscular (TAWA et al., 1997).
—————————-
Nos próximos posts, discutiremos alguns mecanismos moleculares envolvidos no processo de hipertrofia muscular.
Lucas Guimarães
O sistema proteolítico dependente de cálcio foi descoberto na década de 60 (GUROFF, 1967) em neurônios de ratos, e identificado, posteriormente, em outros tecidos, dentre os quais o músculo esquelético (DAYTON et al., 1981; WHEELOCK, 1982). Este sistema depende da ação de proteases ativadas por cálcio, denominadas calpaínas (CROALL & DEMARTINO, 1991). Existem ao menos seis isoformas no músculo esquelético, sendo duas isoformas as principais: a µ-calpaína, e a m-calpaína (TIDBALL & SPENCER, 2002). A principal diferença entre estas isoformas está relacionada à afinidade de ativação pelo íon cálcio. Enquanto que a primeira (também denominada calpaína I) apresenta Km (concentração no qual a atividade é 50% da máxima) entre 5 a 50 µM de cálcio, a segunda (também denominada calpaína II) apresenta Km entre 200 a 1000 µM de cálcio (INOMATA et al., 1983, CROALL & DEMARTINO, 1991).
A ação proteolítica exercida pelas calpaínas parece ser iniciada por um processo de autólise da enzima, que acontece quando esta é exposta a concentrações suficientes de cálcio. Após a remoção de fragmentos de ambas as subunidades das calpaínas, estas apresentam um aumento da sensibilidade a este íon, tornando-se ativas (SAIDO et al., 1994). Entretanto, este mecanismo de ação, confirmado in vitro, é controverso quanto à sua importância in vivo (CARAFOLI & MOLINARI, 1998).
De forma contrária à ação das calpaínas, existe uma proteína, chamada calpastatina, que previne a ativação da proteólise dependente de cálcio, por inibir a ação das calpaínas (DU et al., 2005; GOLL et al., 2007). Desta forma, a relação calpaínas/calpastatina se mostra um importante fator no controle da taxa proteolítica por este sistema. A superxpressão de uma forma negativa da m-calpaína ou de um domínio inibitório da calpastatina em células musculares L8 diminuiu a degradação protéica em 30 3 63%, respectivamente (HUANG & FORSBERG, 1998). Em outro estudo, a superexpressão do transgene da calpastatina em camundongos provocou uma atenuação da perda de massa muscular no sóleo (~30%), em um protocolo de desuso por 10 dias (TIDBALL & SPENCER, 2002). Estes dados demonstram a participação das proteases ativadas por cálcio na degradação protéica muscular. A inibição foi parcial, pois as calpaínas não se mostram capazes de degradar todas as classes de proteínas sarcoméricas (GOLL et al., 2007), sugerindo a importância de outros sistemas proteolíticos neste processo.
Chris Sharma é um verdadeiro fenômeno da escalada esportiva. Nasceu na Califórnia, EUA, e começou a escalar com 12 anos. Aos 14 venceu o campeonato nacional e já encadenou uma 5.14c (Necessary Evil). Aos 15 foi segundo colocado na copa do mundo de escalada, e já encadenava as vias mais difíceis dos EUA.
Assista o trailer e confira as imagens de tirar o fôlego:
[youtube http://www.youtube.com/watch?v=3vEJS2AwGA8&hl=en&fs=1&rel=0&color1=0x006699&color2=0x54abd6&border=1]
INTRODUÇÃO
O músculo esquelético tem sua forma e função altera em resposta a diversos estímulos que modificam a atividade contrátil (exercício, estimulação elétrica, desnervação), carga imposta sobre os músculos (exercícios com sobrecarga, microgravidade), suprimento de substratos (intervenções nutricionais) ou fatores externos como hipóxia e estresse térmico (FLUCK & HOPPELER, 2003). Os avanços nas técnicas de biologia molecular têm permitido a melhor compreensão dos mecanismos celulares e moleculares da plasticidade muscular, ou seja, de como este tecido se adapta às diferentes demandas impostas a ele (GOLDSPINK, 2003).
Figura 01: Estrutura do músculo esquelético
MECANISMOS DE PROTEÓLISE MUSCULAR
O Sistema Lisossomal
O lisossomo foi descoberto na década de 50, identificado primeiramente no fígado de ratos como uma estrutura vacuolar contendo enzimas hidrolíticas que atuam em pH ácido (CIECHANOVER, 2005). O sistema lisossomal contribui de forma importante para a proteólise em diversos processos celulares, tais como turnover de proteínas da membrana celular e degradação de ribossomos (JEFERSON et al., 2001). No músculo, diversas proteases lisossomais são expressas, como as catepsinas B, H, L e D, indicando que este sistema está envolvido na proteólise muscular (JEFERSON et al., 2001). Durante o jejum, os lisossomos contribuem para a taxa geral de proteólise no músculo esquelético, aumentando, particularmente, a degradação de proteínas não miofibrilares. Neste estado, a expressão do mRNA e a atividade da catepsina D encontram-se elevados (JEFERSON et al., 2001). Entretanto, o sistema lisossomal não parece ser a via principal para degradação de proteínas miofibrilares.
———————–
No próximo post continuaremos discutindo os mecanismos de proteólise muscular, com os sistemas ativado por cálcio e ubiquitina-proteassoma.
Lucas Guimarães.
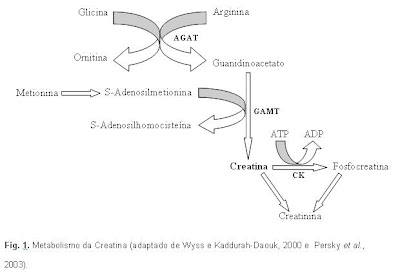


Artigo completo publicado na Revista de Nutrição e disponível em:
Bons treinos!
Lucas Guimarães